
Abstract
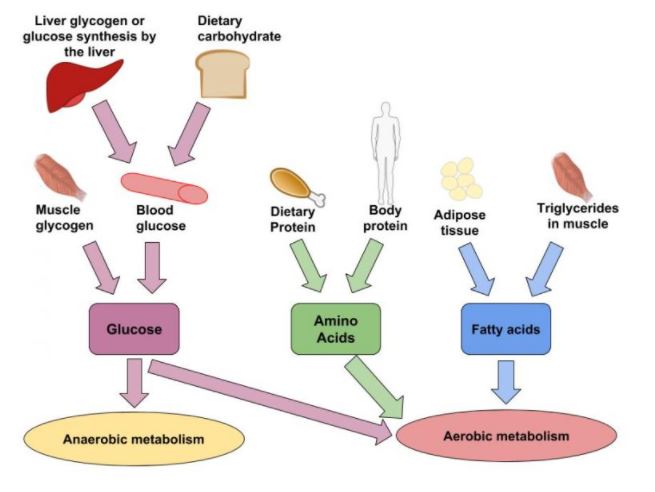
Carbohydrates are an important source of energy for prolonged exercise. During brief intense exercise, it may be the only source of energy for the working muscle and may be derived exclusively from glycogen stores within the muscle fibres themselves. During prolonged submaximal exercise, the magnitude of the contribution that carbohydrates make to the total fuel consumed depends on a number of factors, including (1) exercise intensity, (2) exercise duration, (3) state or type of training the individual has engaged in, or both, and (4) diet previously consumed. Due to their limited storage in the body and the relative ease with which these reserves can be manipulated by combinations of diet and/or training, knowledge of the dynamics of carbohydrate metabolism is an important component in the arsenal of knowledge for coaches and athletes.
Methods
Six healthy, recreationally active men were recruited to participate in this study. Mean ± SE age, body mass, and maximal oxygen uptake (VO2) of the subjects were 28 ± 1 years, 80.0 ± 1.8 kg, and 49.8 ± 1.1 mL kg−1 min. −1 (3.98 ± 0.10 l/min). ), respectively. The subjects were informed verbally and in writing of the purpose of the study, as well as the procedures and risks involved with the experiment. The experimental protocol was approved by the Ethics Committee of Copenhagen and Frederiksberg in Denmark. The study was conducted under the guidelines of the Declaration of Helsinki.
- Blood test.
Heparinized syringes were used to collect blood samples to measure blood Pco2 and Po2 (ABL5, Radiometer), haemoglobin, oxygen saturation (OSM3 hemoximeter, Radiometer), and hematocrit. Arterial and venous plasma samples from heparinized syringes were used to analyze amino acids by high-performance liquid chromatography ultraviolet detection and ammonia by a fluorometric method. Arterial blood samples were collected in tubes containing EDTA and glutathione, which were centrifuged and plasma stored at -40°C for analysis of insulin (DAKO), glucagon (Linco Research) and catecholamines.
Catecholamines were analyzed by high-performance liquid chromatography with electrochemical detection as described by Hallman et al. Arterial and venous samples were collected for glucose, lactate and FFA analysis, measuring blood glucose and lactate on an automatic analyzer (EML105, Radiometer) and plasma lactate and glucose were analyzed with an enzymatic method on an automatic analyzer (EML105, Radiometer). automatic (Cobas Fara, Roche). Plasma FFAs were measured using a WAKO NEFA-C kit (Wako Chemical).
- Muscle analysis.
Biopsy samples were snap-frozen (<10 s) and stored in liquid nitrogen. A 10- to 20-mg muscle sample was used to measure the activity of PDH modified by Putman et al. These data were corrected for differences in blood and connective tissue by adjusting, within a subject, for the highest total creatine concentration in the neutralized perchloric acid extracts of wet muscle homogenates used for PDH activity analyses.
Another part of the biopsy sample was lyophilized, powdered and stored at -80°C. Lyophilized muscle was then extracted with 0.5 M perchloric acid (with 1 mM EDTA) and neutralized with 2.2 M KHCO3. The extract was then used to measure muscle amino acids, glycogen, pyruvate and acetyl-CoA. Adenine nucleotides, phosphocreatine, and creatine were also measured and are reported elsewhere.
Other sources of energy supply
- Glycerol
Glycerol is a hydrophilic sugar alcohol carbon skeleton that can be readily transported as a free molecule in the bloodstream in response to triglyceride hydrolysis and FA mobilization. Lipolytic activity within adipose tissue and intramuscular fat are highly active during moderate-intensity endurance exercise by the action of catecholamines, leading to elevated levels of circulating glycerol.
Glycerol cannot be metabolized by skeletal muscles (since they lack the enzyme glycerol kinase). However, glycerol is transported to the liver via the blood, where it is phosphorylated by glycerol kinase, providing a precursor for gluconeogenesis to help the liver maintain glucose production as glycogen levels decline.
- Ketones
Ketone bodies are low molecular weight, hydrophilic molecules derived from lipids and are normally found in circulation in relatively small amounts ranging from 0.1 to 0.5 mmol/L. The liver synthesizes them in the mitochondria through a metabolic process known as ketogenesis. The central precursor of the ketogenic process is acetyl CoA, derived predominantly from excessive fatty acid oxidation, which in turn is converted to acetoacetate, β-hydroxybutyrate, and acetone.
In order to be used as a metabolic substrate for energy expenditure, ketone bodies are excreted by the liver into the bloodstream and reconverted to acetyl-CoA in extrahepatic organs, and are then incorporated into the TCA cycle for terminal oxidation. Therefore, ketone bodies are widely recognized as an alternative source of energy for highly aerobic metabolic organs such as the brain, skeletal muscle, and heart, particularly when carbohydrate availability is poor or during periods of negative energy balance accounting for approximately 185 g/day as seen in starvation and fasting states, low-carbohydrate, high-fat ketogenic diets, or during prolonged endurance exercise.
The metabolic contribution of ketone bodies as potent metabolic fuel for energy expenditure during exercise and physical activity is an area of increasing attention in the field of exercise physiology and sports nutrition in recent years. However, recent evidence questions the potential ergogenic benefits of low-carbohydrate ketogenic diets and exogenous ketone supplementation in endurance exercise and potential negative health consequences.